Structural Biology
To understand cellular processes on a molecular level the 3D structures of the respective proteins have to be known. NMR spectroscopy is a versatile technique both for elaborate structure determination and for fast interaction studies e.g. for ligand screening. We have studied a diverse range of proteins with the aim to understand how protein function is related to structure.
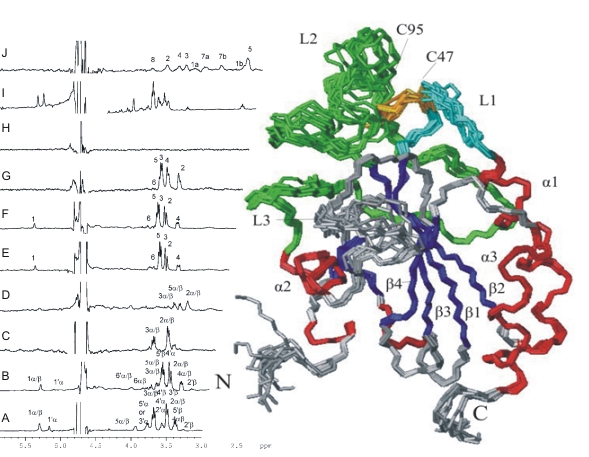
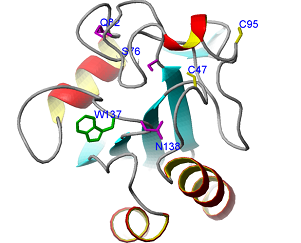
We have solved the structure of a peroxidase from Trypanosoma brucei, the causative agent of African sleeping sickness, and have found that a loop close to the catalytic Cys47 undergoes structural fluctuations questioning the predicted catalytic mechanism. In one of the rare cases, where structure determination has yielded the crucial hints for function, we have identified a novel protein that specifically recognizes N-glycan intermediates during protein glycosylation in the endoplasmic reticulum (ER). It is conceivable that malectin acts as a cochaperone or is involved in the quality control of misfolded proteins. In several studies on domains of the giant muscle protein titin we have investigated titin’s molecular scaffold properties by which it organises the protein patterns of the muscle sarcomer. Another focus has been on the dynamic properties of titin that acts like an elastic, reversing striated-muscle sarcomere stretching.
Membrane proteins still represent a considerable challenge for many structural methods. They adopt a defined three-dimensional structure only in the presence of membrane mimetics, detergents or isotropic bicells. Since membrane protein structure and dynamics often strongly depend on the lipid environment, results obtained in membrane mimetics need to be compared to those in lipidic systems. This is done in collaboration with solid state NMR groups (D. Huster, university of Leipzig, A. Ulrich, IBG 2, KIT). Examples of membrane peptides that we have studied are the antimicrobial peptide sakacin P, the viral oncoprotein E5, the signal peptide PhoD, the membrane anchor of pestivirus glycoprotein E(rns), and the transmembrane domain of the PDGFRβ, a member of the cell-surface receptor tyrosine kinase family.
Apart from structure determination we study in close collaboration with the Cato group at ITG, KIT how mutations affect the structural integrity of Bag1 or the interaction with ligands. Another topic is the identification and analysis of natural compounds isolated from microorganisms in collaboration with the Syldatk group at TEBI, KIT or of peptide mimetica potentially useful in therapeutics.
In collaboration with the Bräse group at IOC, KIT we have studied the conformation of peptoids, protease resistant peptide mimetica. Since conventional 2D NMR did not yield enough constraints for structure determination we have adopted a computational approach to include chemical shifts as structural constraints. Chemical shifts are among the NMR parameters that change most upon structural changes. Structures have been refined against theoretically chemical shifts that have been calculated on the basis of bond polarization theory implemented in the program COSMOS.
Structure gallery
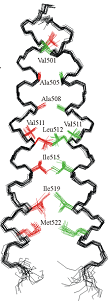 |
Hydrophobic matching controls the tilt and stability of the dimeric platelet-derived growth factor receptor (PDGFR) β transmembrane segment.
PDB-ID: 2L6W |
 |
Malectin: a novel carbohydrate-binding protein of the endoplasmic reticulum and a candidate player in the early steps of protein N-glycosylation.
|
 |
Structural basis for a distinct catalytic mechanism in Trypanosoma brucei tryparedoxin peroxidase.
|
.jpg) |
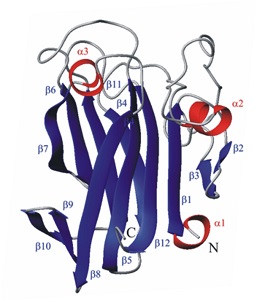
The structure of the FnIII Tandem A77-A78 points to a periodically conserved architecture in the myosin-binding region of titin.
PDB-ID: 3LPW |
.jpg) |
The Ig doublet Z1Z2: a model system for the hybrid analysis of conformational dynamics in Ig tandems from titin.
PDB-ID: 2A28 |
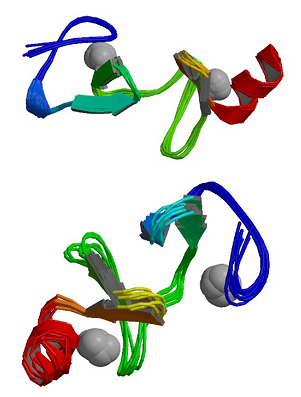 |
Structure and dynamics of the human muscle LIM protein.
|
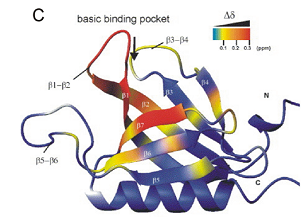 |
Structure and phosphatidylinositol-(3,4)-bisphosphate binding of the C-terminal PH domain of human pleckstrin.
PDB-ID: 1XX0 |
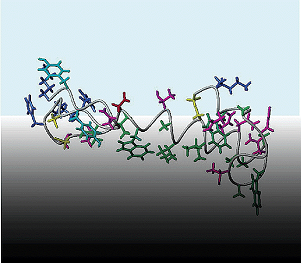 |
Three-dimensional structure in lipid micelles of the pediocin-like antimicrobial peptide sakacin P and a sakacin P variant that is structurally stabilized by an inserted C-terminal disulfide bridge.
PDB-ID: 1OG7, 1OHM, 1OHN |
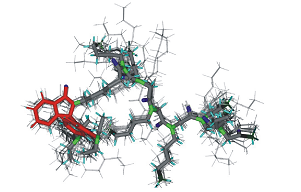 |
Structural characterization of a peptoid with lysine-like side chains and biological activity using NMR and computational methods. Pubmed |